
请教八十一子,这些理论我看着很make sense,只是hypothesis吗?
还有关于分子结构的非对称,除了极个别的,生物体内都是氨基酸左旋,糖右旋,虽然实验室合成的两个方向都可能。这个能给讲讲为什么吗?
============
浮生, 当你说到重复性和对称性时, 我一下子想到动物世界,是如何从一个圆圆的细胞长成各个不同的种属型状, 但它们又多拥有共性, 如前后, 头尾,中轴。
复习了一下胚胎学, 所有的animal architecture都具有这样的共性: modularity and pattern repetition,symmetry and polarity,从低等动物如starfish 到人都encoded在胚胎的发育中。
人的那种对这些建筑in awe的情感, 是否来自与机体深埋的同样的机理?
现在我要上班, 呆会看能否上些图, pictures worth thousand words


 手的x-ray, 是说明系列的reiterated 构建,不同的只是shape和number。
手的x-ray, 是说明系列的reiterated 构建,不同的只是shape和number。
 最后一个彩图是repetition in Fine scale: 这是放大的蝴蝶翅膀的花纹,每个小的unit是细胞level,每个细胞是一个single scale的颜色,collectively, 它们组成一个几何图案,和你及八爷谈的马赛克就是一回事。
最后一个彩图是repetition in Fine scale: 这是放大的蝴蝶翅膀的花纹,每个小的unit是细胞level,每个细胞是一个single scale的颜色,collectively, 它们组成一个几何图案,和你及八爷谈的马赛克就是一回事。
============
ZT: Evolutionary Theories of Asymmetrization of Organisms, Brain and Body (I)
Features of the symmetry are determined by the isotropy of the environment. Maximal extent of organism symmetry corresponds to a completely isotropic ecological niche.
First organisms on Earth floating in the depths of water unicellular and lower multicellular organisms had the maximum possible spherical symmetry. They appeared approximately 3.5 billon years ago. Anisotropic environment step by step was leading to asymmetry of the organisms.
Asymmetrization along the “top – bottom” axis occurred under the influence of gravity. This led to the appearance of the attached, low-mobile forms (plants and coelenterates) that had radial symmetry.
Asymmetrization along the “front – back” axis occurred due to the interaction with space, when rapid motion was required (to escape from the predator, or to chase a pray). As a result, the main receptors and the brain were moved to the front of the body. Organisms with bilateral symmetry were dominating last 650-800 million years. These are crustaceans, fish as well as the most progressive forms, i.e., mammals, birds, and insects.
V. Beklemishev (1964) distinguished three types of symmetry (spherical, radial, and bilateral) and arranged them in evolutionary array (Fig. 1). Forth type of triaxial asymmetry (aaa) he mistakenly assigned to a primitive organism (amoeba) and placed at the beginning of the array. Organisms of bilateral symmetry were considered the “crown” of evolution.
Amoeba → spherical → radial → bilateral symmetry
 Fig. 1. The evolutionary array of organisms’ symmetry types.
Fig. 1. The evolutionary array of organisms’ symmetry types.
c — symmetry, а — asymmetry of an organism on
each of the 3 axis of the three-dimensional space.
What determines asymmetry along the “left – right” axis was unknown.
Basic statements of the new theory:
1. The evolution of any structure goes from symmetry to asymmetry. The asymmetry of the organisms in the phylogenesis grows on all three axes (I — dorso-ventral (back – belly), II — anterior-posterior (nose – tail), III — and mediolateral (left – right).
2. Symmetry and asymmetry are features of a shape. Amoeba is shapeless and therefore should be removed from the evolutionary array. Extrapolation of evolutionary logic of the existing array: ccc ® аcc ® ааc have allowed to state a hypothesis about further asymmetrization of modern progressive forms and their natural transition from bilateral symmetry to the last type—triaxial asymmetry (TA) (аас ® ааа). Growing number of the facts of lateral asymmetry found in modern progressive forms (functional asymmetry of a brain, right handedness in humans, unilateral ovulation and unihemispheric sleep of dolphins) confirms this transition.
3. Which factor of the environment dictates the development of lateral asymmetry?
According to the idea of asynchronous evolution such factor can be time, so one side (organ) is more advanced ("vanguard" as, already in the future) and another one is a "rear-guard" (as yet in the past). Triaxial asymmetry type should be the most evolutionary advanced (Fig. 3).
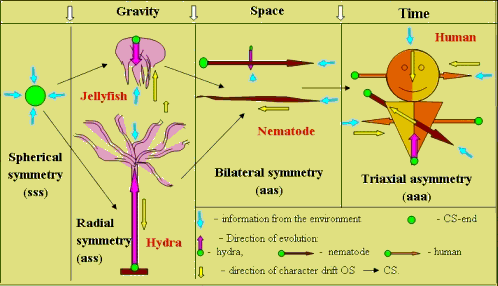 Fig. 3. Adaptive asymmetrization of organisms in anisotropic environment
Fig. 3. Adaptive asymmetrization of organisms in anisotropic environment
4. So, the information flows from the isotropic environment create asymmetry in organisms. How the field acts upon the organism and why it evolves? Let’s take one-dimensional (linear) organism — hydra in the field of gravity and earthworm (or snake) in space.
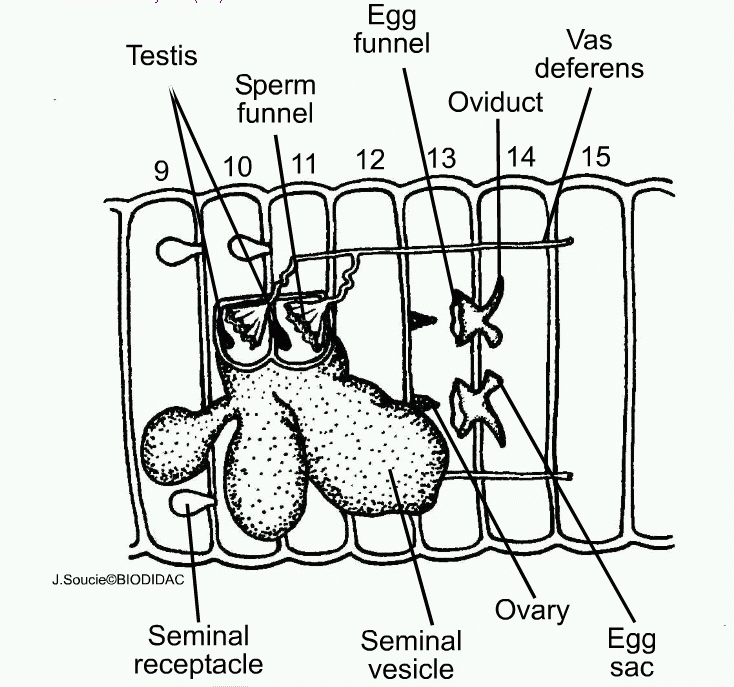
Phenotype appears from interrelation of genetic and environment information: P = G + E. The source of genetic information is zygote. Since the organism is developed from the zygote, in hydra it should be located at the bottom of the foot, and information flow should move to the top towards tentacles and at earthworm zygote at the end of the tail and genetic information flows to the nose, so tentacles are younger than the leg and nose — younger than the tail. Can development start from the nose? No, because ontogeny repeats phylogeny. Can it start in the middle of linear organism? Yes, if development starts at the border of two environments (ground – atmosphere as in the case of plants, then seed is growing up and down). Information from the environment always go towards (growth) and forms the gradient of potential. Then the new organ (character) should appear at the point of growth. If they are necessary their, like receptors or brain, they stay others drift along the gradient towards the tail. The Evolutionary Theory of Sex allows verifying this hypothesis. Hydra had all three types of reproduction: asexual (using kidneys), hermaphrodite, and dioecious, and earthworm had asexual and hermaphrodite. Theory of Sex predicts that ovaries at hydra should be between kidneys and testes, and at earthworm — closer to the tail than testes. That is so. Hydra has kidneys at the bottom, ovaries higher and testes even higher. Ovaries of earthworm are in the 13th segment and testes — only in the 10th.
5. Organisms of triaxial asymmetry keep two previous asymmetries. Lateral asymmetry appears on a background of previous ones (a ‘back–belly’ from a jellyfish, and ‘nose-tail’ from the opossum), therefore it should spread from front to back.
6. Increase in asymmetry can be seen in evolution of many systems:
The trend towards asymmetry can be followed in phylogeny of plant organs (flower, leaf, fruits, and seeds). It is known, that zygomorphic (bilateral symmetry) flowers [Gladiolus sp., Orchids, Eyebrights and Violets] are evolutionary more progressive, than actinomorphic (radial symmetry) flowers [Primula, Narcissus, Pyrola], but are less progressive, than triaxial asymmetric ones [Cannaceae and Valerianaceae].
The morphology of a leaf during evolution follows the same picture: spherical symmetry of chlorella, radial symmetry of pine needles, bilateral symmetry of Magnolia leafs, and triaxial asymmetry of Begonia or Elms leafs.
The same trend can be found in embryogenesis—spherical zygote, radial gastrula, bilateral embryo and triaxial asymmetric child. Vertebrate embryos (and indeed, many invertebrates) exhibit a strikingly conserved left-right asymmetry of the internal organs. Nearly all visceral organs of the thorax and abdomen are left-right asymmetrical in their anatomy, placement, and, in some cases, physiology. Directional left-right asymmetry is conserved throughout chordate evolution, although the details of anatomical asymmetry can vary among species.
One can conclude that asymmetrization is not especially human phenomenon, but general evolutionary phenomenon inherent to all live systems.
Narcissus, Orchid, Cannaceae



Chlorella, Pineneedles, Magnolia leaf, Begonia









